Карякин И.В., Государственный природный биосферный заповедник «Керженский», г.Н.Новгород, E-mail: Ikar_research@mail.ru
 Динамика численности видов в модели хищник-жертва издавна привлекала исследователей, однако до последнего времени все сводилось к ее анализу во времени. С развитием геоинформационных технологий стал возможен и пространственный анализ данной модели.
Динамика численности видов в модели хищник-жертва издавна привлекала исследователей, однако до последнего времени все сводилось к ее анализу во времени. С развитием геоинформационных технологий стал возможен и пространственный анализ данной модели.
В рамках проекта по инвентаризации биоты и созданию ГИС биосферного резервата «Нижегородское Заволжье» (Нижегородская область, Россия) для территории Керженского заповедника (рис. 1) предпринята попытка анализа распределения и динамики численности хищников и их жертв на примере модельной группы взаимодействующих друг с другом видов: жертвы – заяц-беляк (Lepus timidus), глухарь (Tetrao urogallus), тетерев (Tetrao tetrix), рябчик (Tetrastes bonasia); хищники – рысь (Felis lynx) и беркут (Aquila chrysaetos). В данной модельной группе хищники, имеющие разную стратегию и способы кормодобычи, специализируются на одних и тех же видах (куриные птицы и заяц-беляк), причем зайцы составляют до 70-80% рациона обоих видов хищников на данной территории. В то же время виды-жертвы имеют разную биотопическую приуроченность и флуктуации их численности асинхронны, что позволяет при выявлении тех или иных закономерностей в пространственном распределении говорить о влиянии одних видов на другие внутри модельной группы.
Рис. 1. Керженский заповедник на мозаике снимков Landsat 7.
В качестве основы для анализа были использованы тематические векторные слои, в том числе план лесонасаждений территории заповедника 1989г. и карта основных типов растительности, подготовленная в результате классификации космических снимков Landsat 7 ETM+ 1999-2001гг. В качестве тематических данных использовались предварительно обработанные материалы зимних маршрутных учетов (для рыси, зайца-беляка и куриных), учетов на трансектах осенью и весной (для куриных) и всесезонных маршрутных учетов хищных птиц (беркут). База данных всей совокупности регистраций видов была интегрирована в среду ArcView 3.х, где по каждому виду созданы точечные векторные темы распределения видов. На их основе с помощью дополнительного модуля Spatial Analyst методом Kernel в формате GRID построены карты плотности видов-жертв (рис. 2) и итоговая карта плотности всех видов-жертв. Карта распределения индивидуальных участков рыси построена по совокупности регистраций одиночных особей и выводков в год с помощью модуля Animal Movement 2.0 под ArcView (разработка USGS, см. http://www.absc.usgs.gov/glba/gistools/). Для определения схемы распределения гнездовых участков беркута использован метод построения симметричных полигонов заданной площади внутри контура области возможного гнездования вида. Площадь полигонов гнездовых участков определялась, исходя из среднего расстояния между гнездами соседствующих пар, а область возможного обитания выделена на основании параметров распределения известных гнезд по отношению к тем или иным выделам карты растительности [1].
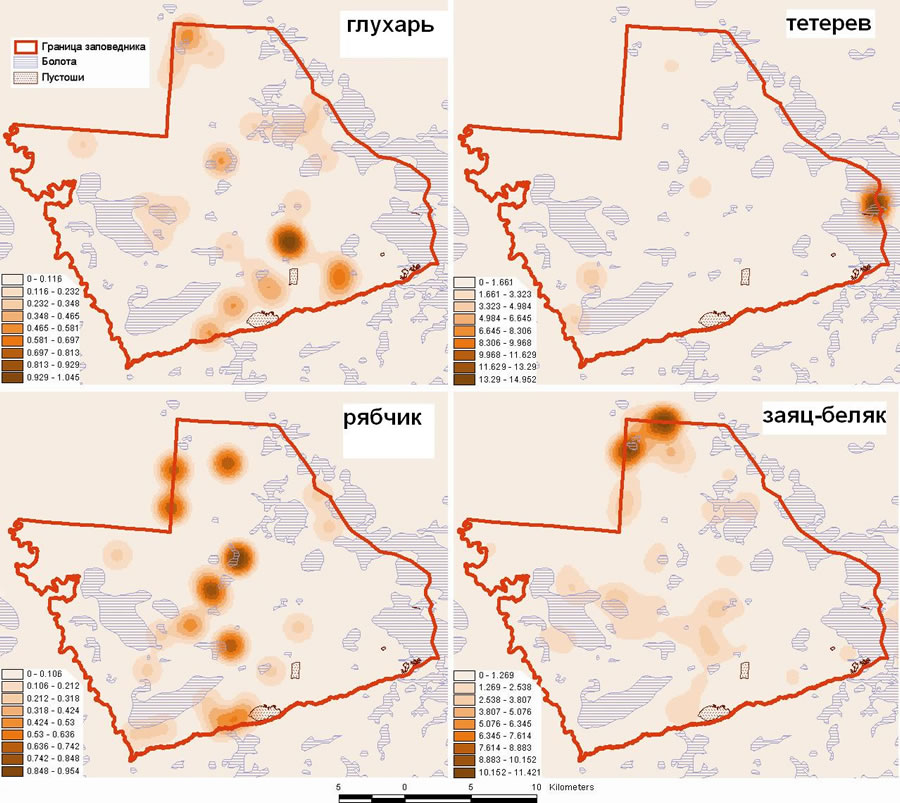
Рис. 2. Карты плотности видов-жертв в Керженском заповеднике в зимний период 2006г.
Сопоставление карт распределения каждого вида по годам и сезонам позволило выявить довольно сильную временную и пространственную динамику у куриных птиц. С 1993 по 2006 гг. их численность изменялась более чем в 10 раз, но районами концентрации оставались местообитания, оптимальные для каждого вида: для рябчика – приречные хвойно-широколиственные леса, для тетерева – комплексы болот, для глухаря – боры. Численность основных объектов питания хищников – зайца и тетерева, как и предполагалось, изменялась асинхронно. Последние 3 года (2004-2006 гг.) численность тетерева в заповеднике росла, но ее динамика в разных очагах, сосредоточенных вокруг токов, приуроченных к крупным болотам, была разной, причем не только позитивной, но и негативной. В то же время численность зайца за этот же период устойчиво сокращалась за счет быстрой деградации группировок, сосредоточенных в периферийной части крупных болотных массивов и сосновых пустошей (рис. 3). Количество индивидуальных участков рыси на территории заповедника в последние несколько лет, по-видимому, остается стабильным. Рысь в разные годы регистрируется на 4-5-ти участках, а самки с выводками – на 2-3-х участках. Количество же гнездовых участков беркута определенно выросло в период пика численности зайца в 2002-2004гг. на фоне реализации мероприятий по устройству искусственных гнезд и в настоящее время является оптимальным для таежной зоны. Территорию заповедника захватывают 7 участков беркута, на 4-х из которых обнаружены гнезда и еще на 3-х зарегистрированы встречи птиц. Сложившаяся структура гнездовой группировки беркута в районе Керженского заповедника в настоящее время остается без изменений, изменяется лишь успех размножения.
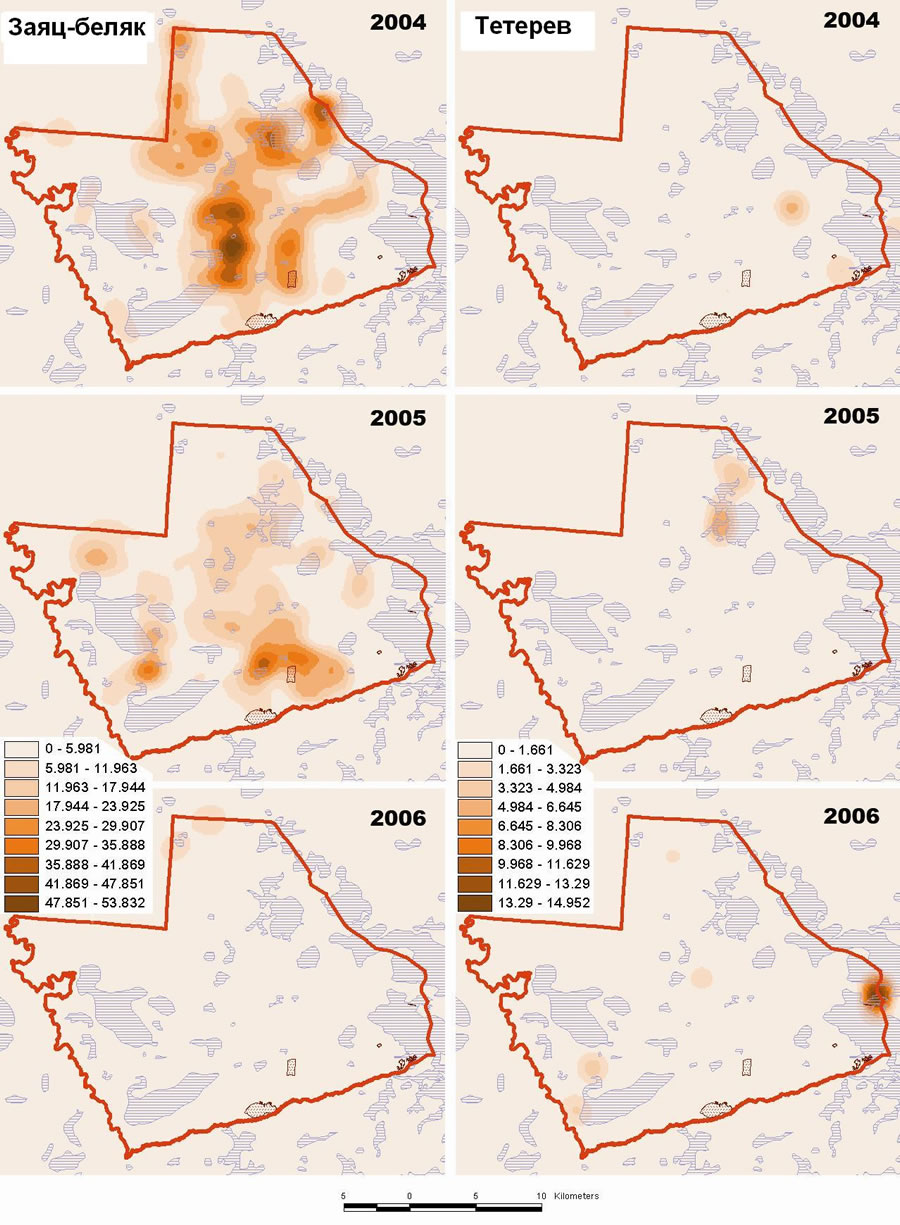
Рис. 3. Динамика распределения и плотности зайца-беляка (слева) и тетерева (справа) в зимний период 2004-2006гг.
Статистический анализ изменений численности видов-жертв в заповеднике по отношению к ряду абиотических и биотических факторов показал значимую корреляцию лишь между плотностью рябчика и количеством осадков [2]. В связи с этим, было высказано предположение о наиболее важном влиянии на численность видов-жертв пресса хищников.
Совмещение итоговых карт распределения и плотности видов-жертв и карт индивидуальных участков видов-хищников (рис. 4) показало высокую степень положительной корреляции между увеличением плотности видов-жертв и удалением от центров индивидуальных участков хищников. В итоге выяснено, что в осенне-зимний период максимальной численности виды-жертвы достигают в оптимальных для обитания биотопах за пределами участков хищников, а в центре их участков плотность жертв в аналогичных биотопах минимальна.
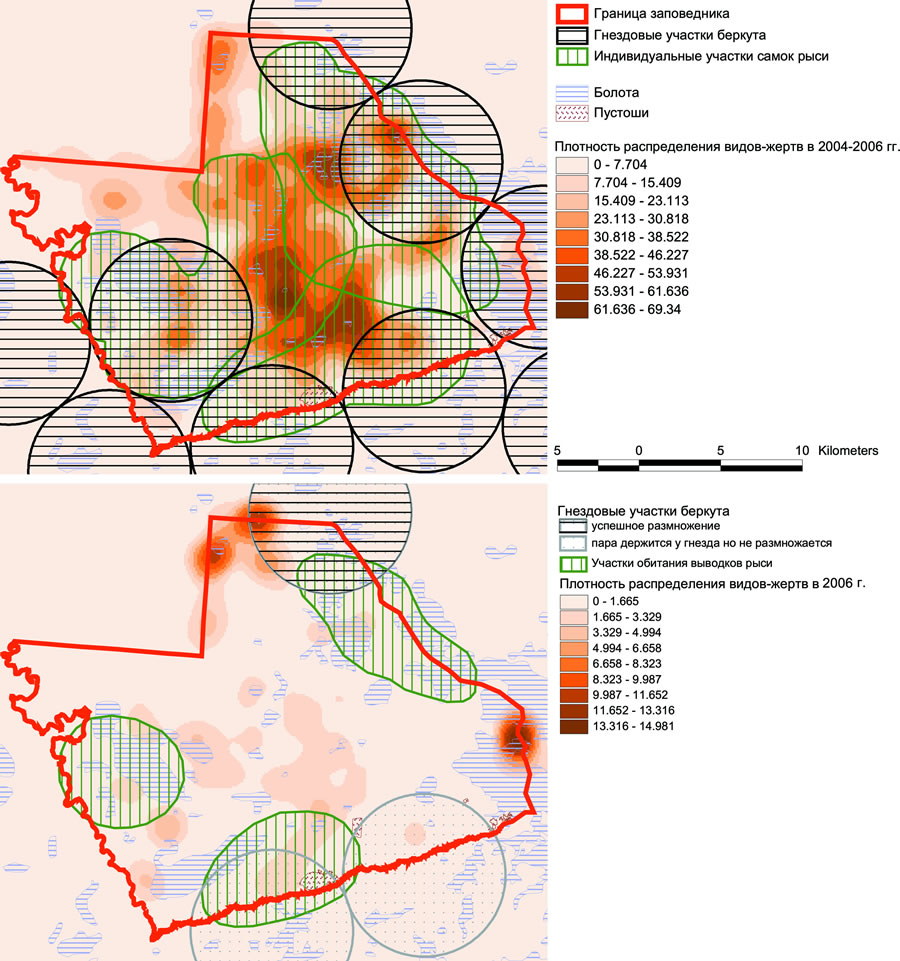
Рис. 4. Плотность видов-жертв и распределение индивидуальных участков хищников в 2004-2006гг. (вверху); плотность видов-жертв и распределение выводков рыси в зимний период 2006г. и посещавшихся гнездовых участков беркута в весенний период 2007г. (внизу).
Индивидуальные участки беркута и рыси постоянны и имеют крупные размеры. На основе проведенного анализа можно предположить, что их пространственная структура в первую очередь определяется биотопическими особенностями местности и лишь во вторую – численностью видов-жертв. Численность видов-жертв в первую очередь влияет на успех размножения хищников. Беркут в своем распространении тяготеет не к местам наиболее плотного обитания какого-либо вида-жертвы, а к экотонным сообществам (граница лес/болото) в непосредственной близости от крупных болот. В этих местах плотность практически всех видов-жертв средняя, но их доступность для добычи с воздуха довольно высокая и имеются специфические условия для устройства гнезда. В итоге, в летний период беркут снижает до минимума плотность всех основных видов-жертв, в особенности зайца-беляка, в центре своего гнездового участка, а в зимний период существенно расширяет территорию охоты, оказывая хищнический пресс в периферийных областях своего участка. Именно по причине такого постоянного локального прессинга в отдельные годы виды-жертвы не успевают восстанавливать численность до оптимальной на участках беркута, и орлы вынуждены делать перерыв в размножении из-за невозможности успешного прокорма птенцов. Поэтому очень часто отсутствие успешного размножения у разных пар никак не связано с общим спадом численности какого-либо вида-жертвы в исследуемом районе в целом. Оно является следствием превышения хищнического пресса на локальном участке. Рысь, как и беркут, тяготеет к экотонным сообществам. Но, в отличие от орлов, она имеет более крупные участки, не связана специфическими условиями для устройства логова и, поэтому, в течение всего сезона предпочитает охотиться на территориях оптимальных местообитаний каждого вида-жертвы в отдельности, придерживаясь очагов их численности. В результате, она более равномерно осваивает кормовой ресурс, постепенно перемещаясь в места повышенной плотности видов-жертв, в особенности зайца-беляка. По этой причине на участках обитания рыси не наблюдается резких сезонных падений плотности видов жертв, хотя в совокупности с беркутом на территориях перекрытия их индивидуальных участков они оказывают очень сильный пресс на зайца, особенно при низкой численности другого доступного корма, такого как тетерев. Таким образом, и беркут, и рысь осуществляют отбор видов-жертв на ограниченных по площади территориях, преимущественно экотонных.
Благодаря неравномерности распределения крупных комплексов болот и сосновых пустошей, к которым тяготеют хищники, между ними сохраняются сильно фрагментированные лесные территории, где охота крупных хищников затруднительна или невозможна по ряду причин. Эти территории являются по сути рефугиумами видов-жертв и центрами их восстановления, на что однозначно указывает пространственный анализ за период с 1993 по 2006 гг. В среднем за многолетний период плотность всех видов-жертв максимальна именно на территориях, где прессинг хищников минимален (рис. 4).
Помимо получения интересных научных результатов, пространственный анализ плотности популяций видов-жертв по отношению к индивидуальным участкам хищников позволяет решать многие практические задачи:
- оптимизировать биотехнические мероприятия как по отношению к видам-жертвам (подкормка, устройство галечников и пр.), так и по отношению к хищникам (например, устройство искусственных гнездовий для беркута);
- улучшать качество территориальной охраны (выделение зон покоя в местах наибольшей концентрации того или иного вида за многолетний период);
- регулировать прессинг охоты в охотхозяйствах.
В подобных исследованиях ГИС является удобным аналитическим инструментом, помогающем выявлять общие закономерности и типичные примеры распределения и динамики биоты.
Литература
1. Карякин И.В., Бакка С.В., Новикова Л.М. Применение ГИС для повышения эффективности мероприятий по восстановлению численности беркута на территории биосферного резервата «Нижегородское Заволжье» // Пернатые хищники и их охрана. № 6. 2006. С.16-20. http://ecoclub.nsu.ru/raptors/RC/06/raptors_conservation_2006_6_pages_16_20.pdf
2. Гелашвили Д.Б., Иванова И.О. Связь биоразнообразия заповедника «Керженский» с погодными условиями 1993-2006 гг. // Труды государственного природного биосферного заповедника «Керженский». Т.3. Нижний Новгород, 2006. С. 58-75.